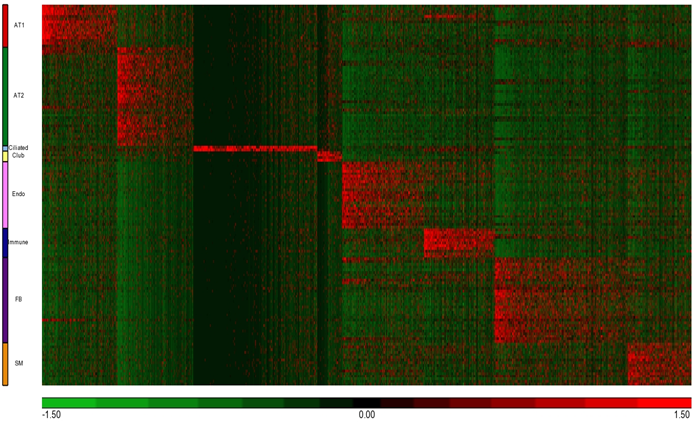
The pulmonary system contains a variety of epithelial cell populations that each reside in distinct anatomical locations. Basal (Krt5, Krt14), secretory, and ciliated cells (Foxj1) line the trachea and the proximal conducting airways; neuroendocrine cells are present in small numbers. The nonciliated, columnar Club cells (Scgb1a1) comprise the majority of the bronchiolar and terminal bronchiolar epithelium in mice, and alveolar type I (AT1: Pdpn, Hopx) and type II (AT2: Sftpb, Sftpc) cells constitute the alveolar epithelium (Bannister, 1999).
The pulmonary system contains a variety of epithelial cell populations that each reside in distinct anatomical locations. Basal (Krt5, Krt14), secretory, and ciliated cells (Foxj1) line the trachea and the proximal conducting airways; neuroendocrine cells are present in small numbers. The nonciliated, columnar Club cells (Scgb1a1) comprise the majority of the bronchiolar and terminal bronchiolar epithelium in mice, and alveolar type I (AT1: Pdpn, Hopx) and type II (AT2: Sftpb, Sftpc) cells constitute the alveolar epithelium (Bannister, 1999).
The pulmonary system contains a variety of epithelial cell populations that each reside in distinct anatomical locations. Basal (Krt5, Krt14), secretory, and ciliated cells (Foxj1) line the trachea and the proximal conducting airways; neuroendocrine cells are present in small numbers. The nonciliated, columnar Club cells (Scgb1a1) comprise the majority of the bronchiolar and terminal bronchiolar epithelium in mice, and alveolar type I (AT1: Pdpn, Hopx) and type II (AT2: Sftpb, Sftpc) cells constitute the alveolar epithelium (Bannister, 1999).
The pulmonary system contains a variety of epithelial cell populations that each reside in distinct anatomical locations. Basal (Krt5, Krt14), secretory, and ciliated cells (Foxj1) line the trachea and the proximal conducting airways; neuroendocrine cells are present in small numbers. The nonciliated, columnar Club cells (Scgb1a1) comprise the majority of the bronchiolar and terminal bronchiolar epithelium in mice, and alveolar type I (AT1: Pdpn, Hopx) and type II (AT2: Sftpb, Sftpc) cells constitute the alveolar epithelium (Bannister, 1999).
'Endothelial cells' (Endo) were readily identified by RNAs defining 'blood vessel development/morphogenesis', as evidenced by the signature genes including: Cav12, Cd34, Fli1, Kdr, Pecam1, Tek, Sox17, and Tie1.
'Myeloid/immune cells' (ML) shared close similarity of 'immune/defense response', 'lysosome', and 'phagocytosis' related RNAs, as evidenced by the signature genes including: Fcgr1/3, Mpeg1, Mrc1, Ly86, and Cyba/b, RNAs expressed by macrophages and monocytes that are not abundant in the normal fetal lung.
The pulmonary lipofibroblast is located in the alveolar interstitium and is recognizable by its characteristic lipid droplets. During alveolar development it participates in the synthesis of extracellular matrix structural proteins, such as collagen and elastin, and as an accessory cell to the type II pneumocyte, in the synthesis of surfactant. (McGowan SE, 1997)
'Matrix fibroblasts' (MFB) expressed RNAs associated with 'extracellular matrix' and 'cell adhesion', as evidenced by the signature genes including: Eln and Vcam, which is a growth factor essential for early lung morphogenesis and alveolarization.
'Myofibroblasts (MyoF)' were identified by RNAs involved in 'actin binding', and 'smooth muscle contraction', as evidenced by the signature genes including: Acta2, Actg2, Myh11, and Tagln.
|